SLIDE 5
- D. T. Scadden, ASH 2012, modified
(data by Li, Frenette, Suda, Morrison, Nilsson, Nakauachi, Nagasawa, Lavesque, Daley, Rafii, Calvi, Adams) Méndez-Ferrer et al. Nature (2010) Carlos López-Larrea et al. Stem Cell Transpl (2012)
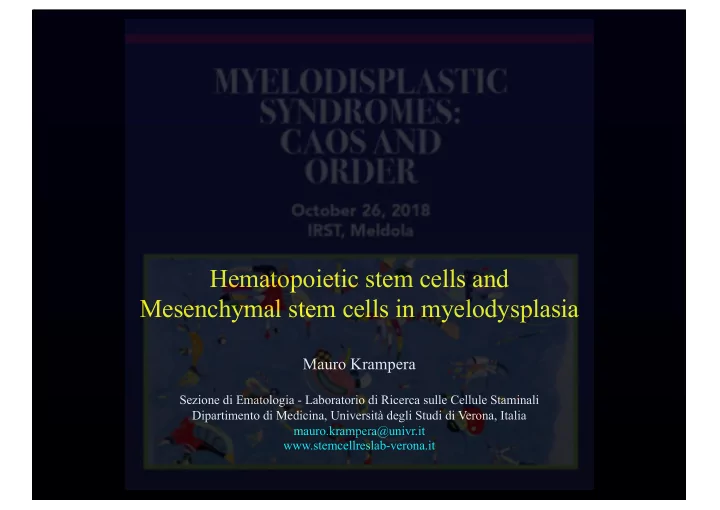
vessel
Endothelial cells
(critical for HSC localization)
Adipocytes
(negatively regulate HSC number)
LeptinR+ MSCs
(source of cKit-L regulating HSC number)
CXCL12+ adventitial reticular cells
(regulate HSC number and localization)
HSCs Nestin+ MSCs
(regulate HSC number and localization)
Osteoblasts
(regulate HSC number and localization)
Bone matrix
(osteopontin limits HSC number Ca2+-R participates in localization) cKit/cKit-L CXCL12/CXCR4 VCAM/a4b1 TGFb1/TGFb-R Ang1/Tie2 BMP-4/BMP-R
Notch1/Jagged1 (+ PTH) Wnt/LRP-Frz
N-Cadherin
Sympathetic neurons
(regulate HSC localization)
Non-myelinating Schwann cells
(regulate HSC quiescence)
Osteoclasts and Macrophages
(regulate HSC localization)
Discrete and specialized micronvironmental space where interactions
through direct contact and soluble factors, amongst: HSCs “Stromal cells” Extracellular bone matrix
(ialuronic acid, glycosaminoglycans,
leading to a finely tuned regulation of HSC functional properties
Bone marrow hematopoietic stem cell niche
Krampera